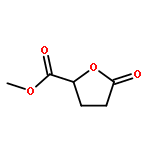
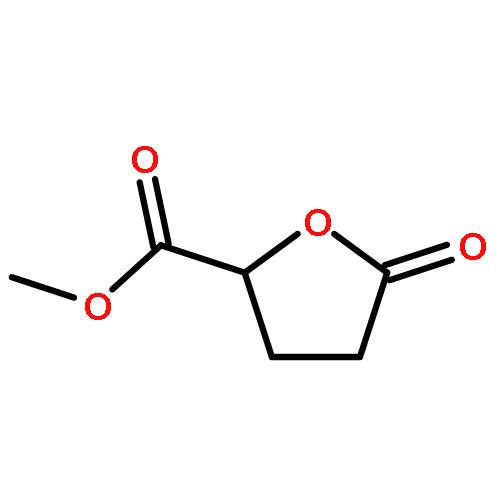
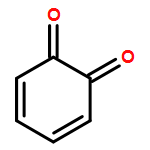
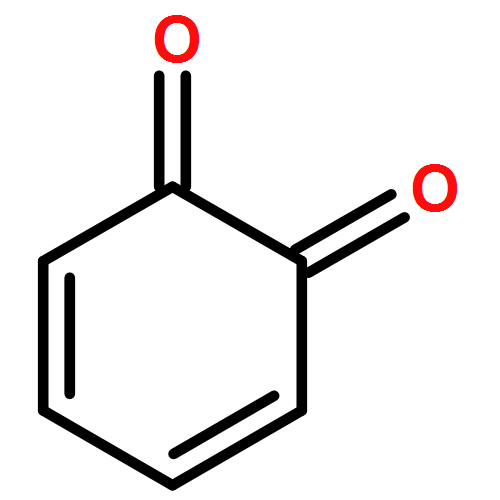
Co-reporter:Ying Ren, Jian Gao, Ji Xu, Wei Ge, Jinghai Li
Particuology 2012 Volume 10(Issue 1) pp:105-116
Publication Date(Web):February 2012
DOI:10.1016/j.partic.2011.02.011
Incorrect folding of proteins in the macromolecular crowding environment in living cells would cause cellular disasters. All cells have developed their structurally and functionally distinct classes of molecular chaperones to help nonnative proteins fold to their native structures, one of which is the most studied GroEL/ES complex. In the present article, large-scale all-atom explicit solvent molecular dynamics (MD) simulations have been carried out on rhodanese folding in a series of chaperonin mutants for 200 ns to understand the mechanism therein. In accordance with experimental results, two factors have been identified to play a significant role, the geometrical confinement effect of the folding cavity and the charge effect of the inner surface of the cavity. Our analysis of the properties during simulation suggests that the GroEL/ES complex directly exerts force on the contacting residues of the substrate, thus assists substrate folding.Large-scale all-atom explicit solvent MD simulations have been carried out on rhodanese folding in a series of chaperonin mutants for 200 ns. Our analysis of the properties during simulation suggests that the GroEL/ES complex directly exerts force on the contacting residues of the substrate, thus assists substrate folding. Architecture of GroEL/ES complex and kinetic movements of residues in rhodanese.
Co-reporter:Ji Xu;XiaoWei Wang;XianFeng He;Wei Ge;JingHai Li
Science Bulletin 2011 Volume 56( Issue 20) pp:2114-2118
Publication Date(Web):2011 July
DOI:10.1007/s11434-011-4543-7
Discrete computer simulations are quite helpful in understanding dynamic structures in complex systems. Recently, using the Mole-8.5 supercomputer and molecular dynamics simulations as a “computational microscope”, we simulated the dynamic structure of a whole H1N1 influenza virion in solution for the first time at the atomic level. In total, 300 million atoms in a periodic cube with an edge length of 148.5 nm were simulated. Using 288 low level hybrids with 1728 C2050 GPUs and a software package developed specifically for the hardware, the simulation executed 770 ps/d with an integration time step of 1 fs, and analyzed the dynamic structure. With the tremendous computational power of GPUs, efficient software packages for various hardware designs, and consistent physical models, more challenging applications will be carried out in the near future.
Co-reporter:Ying Ren, Jian Gao, Wei Ge and Jinghai Li
Industrial & Engineering Chemistry Research 2009 Volume 48(Issue 19) pp:8865-8871
Publication Date(Web):December 12, 2008
DOI:10.1021/ie801441x
The thermal unfolding process of bovine liver rhodanese, composing two globular domains (N-domain and C-domain) with similar tertiary structures, has been studied by explicit solvent molecular dynamics (MD) simulations at high temperatures of 450 and 500 K, as well as 308 K for comparisons. The results are in good agreement with the available experimental results (Horowitz, P. M.; Butler, M. Interactive Intermediates Are Formed During the Urea Unfolding of Rhodanese. J. Biol. Chem. 1993, 268 (4), 2500−2504. Shibatani, T.; Kramer, G.; Hardesty, B.; Horowitz, P. M. Domain Separation Precedes Global Unfolding of Rhodanese. J. Biol. Chem. 1999, 274 (47), 33795−33799. Ybarra, J.; Bhattacharyya, A. M.; Panda, M.; Horowitz, P. M. Active Rhodanese Lacking Nonessential Sulfhydryl Groups Contains an Unstable C-Terminal Domain and Can Be Bound, Inactivated, and Reactivated by GroEL. J. Biol. Chem. 2003, 278 (3), 1693−1699). Besides that our simulation can also depict more dynamic details of the unfolding process. The solvent accessible surface area (SASA) shows a remarkable increase mainly due to a more exposed hydrophobic area, indicating that the hydrophobic interaction is considerably weaker at high temperatures. Comparisons between the thermal stabilities of equivalent secondary structures in both domains suggest that the C-domain is more fragile than the N-domain and the breaking down of the secondary structures follows the pattern [α-helix]→[bend, turn, 3-helix, and 5-helix]. Different regimes of unfolding intermediates have also been discussed.